Lysbuget høgeørn
Changeable Hawk Eagle

Lysbuget høgeørn (Nisaetus cirrhatus)
Taksonomisk klassifikation:
Klasse: Fugle (Aves)
Orden: Rovfugle (Accipitriformes)
Familie: Høgefamilien Accipitridae – 10 underfamilier
Underfamilie: Aquilinae – en af de 10 underfamilier
Slægt: Nisaetus ( 10 arter) – se dem her
Art: Lysbuget høgeørn (Nisaetus cirrhatus)
Den Lysbugede høgeørn forekommer i fem underarter, som er stærkt knyttet til forholdsvis veldefinerede lokationer – se dem HER. Arten på billederne her er fra Sri Lanka, og er derfor underarten “Nisaetus cirrhatus ceylanensis”. Det tredje slægt-navn angiver, at det er Sri Lanka arten (mere om slægtsnavne senere).
Under søgning på denne art stødes der på flere forskellige danske betegnelser som Kamhøgeørn og Forandrelig høgeørn, men det officielle danske navn er ”Lysbuget høgeørn”.
Karakteristisk har de fleste arterne i slægten et par signaturfjer på toppen af hovedet, men dog ikke alle, idet to er kamløse arter, og to underarter af netop den Lysbugede høgeørn heller ikke en kam (signatur fjer), og en underart kun har en meget lille kam eller set ingen (0-5 cm.) – ses listet HER
Lysbuget Høgeørn, skal ikke forveksles med dens nære (og større) slægtning Høgeørnen (Bonelli’s eagle) fra de ægte/egentlige ørne” Aquila-slægten”. Brug af det danske navn “høgeørn” til begge arter kan skabe en del forvirring.
Lysbuget høgeørn hører til en af to slægter af mellemstore ørne (høgeørn-gruppen), som har deres navn fra deres ligheder med høgene – deres afrundede vinger og ensartede lange fjerbeklædte ben. De er typisk langhalede og slanke, egenskaber der giver dem gode muligheder for at jage og færdes i skov-habitater – ganske som de egentlige “høge”..

Den mest udbredte
Blandt medlemmerne af sin slægt ”Nisaetus” (udgøres af 10 arter) skiller den Lysbugede høgeørn sig ud, som den mest udbredte tilpasningsdygtige og artsrigeste art.
Den er en typisk ”høgeørn”, idet den er en adræt skovlevende rovfugl. Som de fleste andre skov-rovfugle har Lysbuget høgeørne (og Nisaetus-arter generelt) en lang hale, korte brede vinger og relativt lange, men kraftige ben. Egenskaber som alle giver fuglene større manøvredygtighed og hurtigere slagtider. De har derved en fysiologi, der gør det nemmere at manøvrerer imellem den tætte vegetation, der findes i deres skovklædte jagtområder, end andre rovfugles krops- og vingestruktur tillader dem.
Disse egenskaber gør det muligt for dem på en effektiv måde, at varierer deres byttevalg mellem fugle, pattedyr eller krybdyr.
Lysbuget høgeørn (og andre høgeørne) er således usædvanlig tilpasningsdygtige, ikke kun efter standarderne for dens slægt, men også efter standarderne for hele dens underfamilie. Mens skovrydning har reduceret populationerne af andre Nisaetus-arter (resulterende i at fire underarter er klassificeret som truede arter), har Lysbuget høgeørn vist sig, at være bemærkelsesværdig modstandsdygtig over for habitatforringelse.
Arten har et udbredelsesområde der strækker sig over tredivetusinde kvadrat-kilometer, og med et revir på omkring 1.200 km. per par, kræver denne ørneart en ganske stor privatsfærer.
Udbredelsesområder indbefatter det Indiske subkontinent, det sydøstlige Asien, Sri Lanke, Nepal, Burma, det sydøstlige Asien m.m.

Farve og variation
De enkelte individer udviser en bred farve-variation i deres fjerdragt, gående fra bleg til mørk-brun chokolade og næsten sort, varierende alt efter fældning og alder, hvilket har givet anledning til det engelske navn ”changeable” (foranderlig) Hawk-eagle.
Det forandrelige består i to farvevarianter (farvemorf) ”bleg og sort” og alle former for mellemliggende variation. Bleg morf voksen er mørkebrun foroven med et hvidt bryst overtrukket med mørke striber. Den unge ørn er meget lysere i det brunlige, med renere hvid underside og mindre omfattende striber overalt på kroppen.
Mørk morf er mørk over det hele med blegere ydre vingefjer og kan forveksles med mørke honningmusvåger; men høgeørnen har meget længere vinger og en mere slank profil.
Hvad er en farvemorf?
Polymorfi fra en (DNA)-sekvens-variation.
I biologien bruges betegnelsen polymorfi om forekomsten af flere udgaver af et gen i en population, der forårsager forekomsten af to eller flere individformer (fx farvevarianter / farvemorfer).
To forskellige farvemorfer (varianter)
Den Lysbugede høgeørn findes i to forskellige former eller farvemorfer. Dette er en almindelig form for polymorfi (poly = mange; morf = form), der forekommer hos fugle, især rovfugle.
Hos den Lysbugede høgeørn er der en bleg morf og en mørk morf, samt mange mellemliggende farver og mønstre. Den voksne blege morf fremstår mørkebrun foroven og hvidlig med mørke striber forneden. Voksen mørk morf er dyb mørkebrune til nærmest sort. Hver morf kan have specifikke fordele i visse habitater, såsom visse overlevelses- og reproduktive fordele.
Arten forekommer også i en variation af den karakteristiske ”kam” (signaturfjer), den har på hovedet. Idet nogle arter er kamløse, andre har en stor kan, og andre igen en mindre. Den engelske betegnelse for art med kam og art uden, kan også være ”crested Hawk-eagle” og ”cresless Hawk-eagle. Mere om det senere.

Billederne her over er af Nisaetus cirrhatus ceylanensis (Sri Lanke underarten) som er monomorf, hvilket angiver at underarten ikke omfatter den helt mørke (nærmest sortlige morf (farve) variant).
Farvepolymorfi (flere farve varianter) - Artskomplekset
Farvepolymorfi forekommer hos omkring 3,5 % af alle fuglearter, men er mere almindelig hos rovfugle og især hos Accipiter-høge, hvor omkring 25 % af arterne er polymorfe (farvevarianter).
Accipiter er en slægt af rovfugle i familien Accipitridae, som omfatter 51 anerkendte arter (fx Spurvehøg), og som er den mest forskelligartede slægt i rovfugle familien, når det drejer sig om farve og mønster. De findes hovedsageligt i skovklædte eller buskede områder.
Farvemorfer findes hovedsaligt hos rovfuglene – især hos ”høgegruppen”
Forskere har længe været fascineret af farvepolymorfi (farvevariation) inden for samme art, som har samme levested (lever i samme biotop), fordi forekomsten af to eller flere fænotyper i samme population er i modstrid med forestillingen om, at selektivt tryk bør favorisere den bedst tilpassede og dermed den optimale form for eksistens i et bestemt miljø.
Farvemorfer påvirker individers præstation gennem direkte effekter af deres pigmentproduktion (f.eks. camouflage eller termoregulering), samt indirekte effekter (f.eks. fysiologiske og adfærdsmæssige træk). Derfor er det usandsynligt, at forskellige morfer er fordelt tilfældigt i miljøet, og snarere sandsyneligt, at én bestemt variation over tid vil komme til at dominere en population på en bestemt lokation.
Forskning har dog vist, at farvevariation (farvepolymorfi) inden for parret, kan give mere føde til parrets unger, hvor fx en mørk morf bringer mere føde på lysfattige dage (samt i det tidlige morgen og sene eftermiddagslys) i forhold til lyse morf (utvivlsomt fordi de falder bedre ind i områdets baggrunds-billede).
Når de to morfer yngler sammen i et “blandet” par, kan de således supplere hinanden, ved samlet at udvide de bedste betingelser for jagt i forhold til – dagslystimer, byttedyr eller habitatstyper.
Ideen om, at par bestående af de to morfer komplementerer hinanden, kaldes “komplementaritets-hypotesen”.
Redekamera-data viser, at par med blandede morf konsekvent lever mere føde til reden end par med forholdsvis ens morf. Dette indikerer, at de to morfer vælger at tilpasse deres jagt under forskellige omgivende lysniveauer og baggrunde, hvilket udvider deres fourageringsniche som et par. Denne forskning blev publiceret i Behavioral Ecology, og er en del af Carina Nebels ph.d.-afhandling, som blev tildelt i 2021.
Nyerere forskning har yderligere fundet, at fuglearter med flere fjerdragt-farve og mønstre inden for den samme population, udvikler sig til nye arter hurtigere end dem med kun én farveform, hvilket bekræfter en 60 år gammel evolutionsteori.
“Nu hvor dette mønster for første gang er identificeret, er næste skridt i forskningsprocessen, at teste nogle af de foreslåede forklaringer på, hvorfor farvepolymorfi fører til accelereret evolution.” men dette er en helt anden historie.

- Chlangable Hawk-eagle SOP. License: CC BY-SA 3.0

- Changable Hawh-eagle. ELJA. License: CC BY-SA 4.0
Pardannelse mellem individer med træk, der er mere forskellige end sandsynligt ved tilfældig parring – en fordel
Naturlig pardannelse mellem farve-polymorfe rovfugle er en mekanisme der opretholder farvepolymorfi (farve-differentiering) i populationer, og som derved bevarer artens mangfoldighed.
Opretholdelse af et sådant parringssystem (artskompleks) kan forekomme, hvis det giver fordele for mangfoldigheden ved at kønnene vælger en modsat kontrasted morf. Eksempelvis nævnes det, at hos den sorte spurvehøg (Accipiter melanoleucus), kan blandede morf-par have en selektiv fordel, fordi de producerer afkom, der har højere overlevelsesrater grundet deres farve- og mønster kompleksitet – afkommet falder ganske enkelt bedre ind i omgivelserne.
To hypoteser, der kan forklare mekanismen, er hypoteserne om “undgåelse-billedet” og “komplementaritet”. Den første tyder på, at byttedyr inden for et rovdyrs territorium udvikler et søgebillede efter den mest almindeligt forekommende forældremorf, for eksempel han-morfen i rugeperioden, hvor hans morf-mønster vil blive byttedyrenes ”søgebillede”.
Hunner med en modsat farve eller mønster i forhold til sin partner vil så have højere fangstrater og hastighed, når hun begynder at jage i den senere rede-fase, hvor der er et forøget behov for øgning af jagtbytte.
Således forudsiger hypotesen om byttets udvikling af et søgebillede højere leveringsrater for par med blandede morf.
Alternativt hævder “komplementaritets-hypotesen”, at forskellige farvemorfer udnytter forskellige miljøforhold, hvilket tillader blandede morf par, at jage under en bredere vifte af lokation og lysfaser, og forudsiger, at maden dermed leveres mere konsekvent.
Carina Nebet mf. testede disse hypoteser ved hjælp af redekameraer-data til at registrere leveringsrater for byttedyr i den sene redefase, hvor begge forældre er på jagt. De fandt støtte til “komplementaritets-hypotesen”, hvor par med blandede morfe leverer mad mere konsekvent, men ikke med en højere hastighed.
Læs Carina Nebel mf. artikkel i Behavioral Ecology: HER



I parringstiden oplyses om følgende adfærd
Lysbuget høgeørn er som mange rovfugle, en stort set ensom levende fugl, men i yngleperioden opholder de sig i et tæt sammenholdende par, og danner ofte et livslangt parforhold. Både han og hun engagerer sig i den territoriale manifesteringer i udkanten af deres yngleområde, for at afvise andre artsfæller i at trænge ind.
Skulle an artsfælle vise sig, påbegyndes en luftopvisning (normalt startende af hannen), men nogle gange vil også hunnen deltage i fremvisninger, som ofte indledes med, at deres vinger og hale bues opad i overdrevne balancer og mønstre.
Under fremvisningen skydes der lodret op afløst med dybe næsedyk eller skarpt bøjede afbrydelser. Hvis den rivaliserende høgeørn fortsætter med at engagere sig, vil de to rivaler med “fuld hastighed” flyve direkte mod hinanden, og lave en komplet loop-the-loop-vending i luften. De kalder ofte højt under hele forløbet.
Rede, Æg og Unger
Både hannen og hunnen af Lysbuget Høgeørn samarbejder om at redebygningen. De bygger en stor stokket-rede i hovedgaflen på et højt træ.
Deres reder kan være ret store og måler mellem 95-105 cm i diameter og 35-120 cm dybe. For at lægge sidste hånd på deres rede, vil de fore den med grønne blade.
Parret vil forblive på deres territorium året rundt, selvom deres redesæson kun varer omkring 4-5 måneder hvert år.
De oplysninger der fremkommer om antallet af de æg der lægges, varierer en del, idet det både angives, at der kun lægges ét enkelt, og nogle steder op til tre æg. Men de oplysninger jeg har set, og som jeg finder mest valide er, at der kun lægges ét æg!
Ægget (eller æggene) er hvidt med rødbrune pletter. Biologer har observeret, at rugepligten næsten udelukkende falder på hunnen. Hvis hannen hjælper, er det kun sjældent, til gengæld har hannen et andet vigtigt job at udføre, da han har ansvaret for at bringe føde til hunnen, mens hun passer ægget/æggene, som klækker omkring 40 dage efter det er lagt (også her kan der findes angivelser på op til 50 dage!).
Efter yderligere 35 dage aftager den tid gradvist, hvor hunnen behøver at varme ungen, og væksten af fjer og kropsstørrelse accelererer. Hunnen er begyndt at deltage i fødeskafningen, og byttet bliver leveret på nærliggende grene, i stedet for direkte til reden, så ungen tvinges til at forlade reden. Når forældrene nærmer sig med et bytte, kalder de, hvilket tilsyneladende tilskynder den unge ørn til at vove sig ud af reden.
Efter ca. 52 dage er ungen fuldvoksen, men flyver ikke ud før omkring den er 60-68 dage. Den samlede rede-afhængighed blev registreret som 81 dage i Indien. Imidlertid blev den samlede ynglecyklus registreret til at vare omkring 112 dage i Vest-Java.

Underarter og artskomplekset af Lysbuget høgeørn
Den Lysbuede høgeørn (Nisaetus cirrhatus) er (som nævnt tidligere) den mest udbredte asiatiske ørneart, med et meget bredt udbredelsesområde (33.155 km2).
De forskellige underarter er fordelt rundt i Asien, lige fra det indiske subkontinent, til Filippinerne – se boks her under.
På grund af deres brede udbredelse kombineret med de meget forskelligartede geografiske forhold der findes i Sydøstasien, har arten dannet 5 underarter; Nisaetus c. cirrhatus, Nisaetus. c. ceylanensis, Nisaetus c. andamanensis, som danner ”kam-gruppen”.
Indonesien huser de to sidste arter Nisaetus c. limnaeetus og Nisaetus c. vanheurni som udgør en ikke-kammet gruppe.
Lysbuget høgeørn findes således i 5 underarter, som hver har sin geografiske udbredelse – men overlap forekommer.
Hvad er forskellen mellem arter og underarter?
1) En underart er en gruppe inden for en art, der er blevet noget fysisk og genetisk lidt anderledes end resten af gruppen.
2) En art er en gruppe levende væsener med meget ens egenskaber, der kan krydse hinanden og producere levedygtigt afkom.
Geografisk udbredelse:
- Nisaetus cirrhatus cirrhatus : Halvøen Indien
- Nisaetus cirrhatus ceylanensis : Sri Lanka
- Nisaetus cirrhatus andamanensis : Andamanøerne (beliggende mellem Indien/Sri Lanka og Tailand)
- Nisaetus limnaeetus limnaeetus : N. Indien til Indokina, Malaya, Sundaøerne og Filippinerne
- Nisaetus limnaeetus vanheurni : Simeulue I. (off w Sumatra)
Nisaetus floris blev indtil for nyligt betragtet som underart til N. c. Cirrhatus, men har opnået annerkendelse som selvstændig art: findes ved mindre sundaøer (Bali, Lombok, Flores og Wetar)
Kilde: Avibase
To relativt forskellige grupper
Ud over den store farvevariation forekommer Lysbuget høgeørn i to relativ forskellige typer af underarter. De fem underarter kan der ud fra opdeles i to grupper; en med kamme og en uden eller med næsten ikke synlige kam (også kaldet signaturfjer).
En undersøgelse fra 2020 viste, at den herskende reproduktive isolation*, som grundet deres store udbredelse findes mellem de to grupper, var svag. Undersøgelsen anbefalede at behandle de to grupper som artsspecifikke. Eksempelvis findes mørke morfer kun hos nogle af populationens medlemmer, det samme er tilfældet med den signifikante ”kam”.
Men en opdeling på artsplan er ikke effektueret.
* Reproduktiv isolation - Artsdannelse.
Artsdannelse er som regel ikke en proces, man umiddelbart kan observere.
Ofte foregår den ved, at en art bliver geografisk opdelt i to adskilte populationer.
De udvikler sig derefter langsomt over mange generationer i forskellig retning, indtil de rent genetisk er blevet adskilt fra hinanden, og ikke længere kan reproducere sig med hinanden.
Dette fænomen kaldes “reproduktiv isolation*” – altså når der opstår en barriere, som forhindrer parring mellem individer af samme art.
Vælger partner efter udseendet
En usædvanlig type af artsdannelse kan foregår inden for en population – altså uden geografisk adskillelse.
Det sker ved, at et bestemt træk, f.eks. et farvemønster, bliver brugt i partnervalg, så hannen kun parrer sig med en hun med samme farve eller mønster som ham selv, på trods af at en parring mellem to individer med forskelligt mønster også ville resultere i levedygtigt afkom.
Sådan et selektivt partnervalg resulterer i reproduktiv isolation, hvor individer inden for samme population kun vælger partner med et bestemt udseende. Dermed kan nye arter opstå via selektivt partnervalg.
Men det modsatte kan også være tilfældet hos populationen, hvor en mørk morf i stor udstrækning foretrækker en lys morf. Herved fremkommer et stor spredning af farve og mønstre hos populationen. Denne form er velkendt hos Lysbuget høgeørn og underarter.

Gruppeopdeling af de fem underarter - nærmere gennemgået
Kammet høgeørn gruppen (crested hawk-eagel group)
Nisaetus cirrhatus:
Den Indiske art, og den nominelle underart (og således “type-arten” for hele slægten), findes på den Indiske halvø, fra Gangetic-sletten sydpå gennem resten af landet. Den er monomorf i farven. Hvilket betyder, at den meget mørke (nærmest sortlige) farvevariant ikke findes hos denne underart. Betegnelsen ”morf” er beskrevet – Teksten kan genlæses HER.
Nisaetus cirrhatus er relativt bleg sammenlignet med andre af slægtens underarter, men med en lidt mere rødbrun nuance i hovedet. Den har også en tendens til at være mindre stribet.
Toppen er generelt også længere hos denne underart, derfor omtales den almindeligvis på engelsk som ”crested høgørn” – kammet høgeørn.
Nisaetus cirrhatus ceylanensis (Sri Lanka arten):
Denne underart er endemisk for Sri Lanka. Den har gennemsnitlig en lidt længere kam end den nominelle underart. Den er monomorf ligesom den nominelle art, og er også gennemsnitlig blegere og mindre stribet, og fremstår således mere lys-brunlig end resten af gruppens underarter.
Men et lignende udseende er blevet bemærket for fugle fra det sydligste Indien (der ligger tæt på Sri Lanka – kun adskilt af vand). De har farve og mønstre, der udviser stor lighed med Sri Lanka varianten. Hvilket grundet den naturlige hybridzone utvivlsomt skyldes hybridisering imellem de to underarter.
Nisaetus cirrhatus andamanensis:
findes på Andaman-øerne (beliggende mellem Indien/Sri Lanka og Tailand). Underarten er mørkere end den nominerede underart og ligner mere Nisaetus c. limnaeetus i nuance, men angives til ikke at se ud til at have en fuld mørk morf. Den anses for at være den største af underarterne.
Arten angives nogle steder til kamløs, men andre steder til at have en lille varierende kam på 0-5 cm. Den anbringes gerne blandt de kam-løse, men jeg har valgt, at placere den i kam-gruppen, grundet det forhold at den er en underart, faldet efter Nisaetus cirrharus, og derfor nærmere beslægtet med den kammede gruppe, på trods af at den kun fremviser en meget lille kam eller slet ingen.
Ikke-kammet Lysbuget høgeørn gruppe (crestless hawk-eagel group)
De ikke kam-bærende underarters polymorfi (DNA-sekvens-variation påvirkende farve- og mønster) er den egentlige kilde til artens engelske navn ”Changable Hawk-eagle” (foranderlig høgeørn). Den blege form af disse underarter anses, at være mørkere end den nominelle art fundet længere sydpå i Indien (Nisaetus cirrhatus cirhatus) – både i deres lyse- og mørke form. Men farvemæssigt spænder de over hele variationen fra lys- til helt mørk morf.
De kan have en tendens til at være større end de kammede arter.
De to grupper er tilsyneladende tæt på at opnå anerkendelse som særskilt art. Ø-taxaen som stammer fra Nisaetus c. limnaeetus ser ud til at have undergået
”grundlæggervirkninger”* (founder effect), hvilket har begrænset deres genetiske mangfoldighed.
I den kontinentale population er den genetiske diversitet betydelig. Men da det evolutionære mønster af to undersøgte gener ikke stemte overens, og prøver af oprindelse heller ikke viste klare strukturer anses arten Nisaetus c. liminaeetus derfor for en metapopulation*.
*En metapopulation består af en gruppe af rumligt adskilte populationer af samme art, som interagerer med hinanden (ved fælles pardannelse i overlapsområder medførende hybrid-afkom med intermedierne ligheder).
Grundlæggereffekt (founder effect)
I populationsgenetik er
grundlæggereffekten tabet af genetisk variation , der opstår, når en ny population etableres af et meget lille antal individer fra en større population. Som et resultat af tabet af genetisk variation kan den nye population være markant anderledes, både genotypisk og fænotypisk, fra den forældrepopulation, hvorfra den er afledt. I ekstreme tilfælde menes grundlæggereffekten at føre til artsdannelse og efterfølgende udvikling af nye arter.
Grundlæggereffekten er et særligt tilfælde af genetisk drift, der opstår, når en lille gruppe i en population splitter sig fra den oprindelige population og danner en ny. Den nye koloni kan have mindre genetisk variation end den oprindelige population, og gennem tilfældig udveksling af alleler under reproduktion fra efterfølgende generationer fortsætter gruppen hurtigt mod fiksering. Denne konsekvens af indavl gør dog kolonien mere sårbar over for udryddelse.
De kamløse underarter
Nisaetus cirrhatus limnaeetus:
Denne underart findes gennem Himalaya-foden fra Nepal, det nordøstlige Indien, via Burma til store dele af Sydøstasien, inklusive den malaysiske halvø langs Wallace Line (i syd; Bali-Lombok og i nord; Borneo-Suawesi) til Filippinerne og de større Sunda-øer.
Den er den mest udbredte af slægtens underarter.
Mørke morf individer har en tendens til at være chokoladebrune over det hele, selvom halebunden kan virke lysere under flugten. Nogle individer har en ikke udviklet kam på kun 1 til 3 cm i længden, men mange ser ud til helt at mangle den.
Nisaetus cirrhatus vanheurni
Arten er hjemmehørende i Simeulue Island (ved Sumatras vestkyst). Den ligner i udseende Nitaetus c. limnaeetus, men mangler tilsyneladende en mørk morf. Underarten har også meget mindre sorte pletter på sit hvidere udseende bryst og bærer aldrig en kam.
Flores-høgeørn ( Nisaetus floris).
Endemisk art som findes på de mindre Sunda-øer i Indonesien. Er medlem af slægten Nisaetus. Er nævnt her fordi den indtil for “nyligt” blev betragtet som en underart af den Lysbugede høgeørn (cirrhatus, Indien-underarten), men genetiske undersøgelser har vist, at den er en selvstændig legitim art – den har ingen kam.
Flores anses også for at være en meget gammel art.
Artsstatus over Nisaetus underarter og artskompleks.
De fem underarter har udviklet sig fra to af slægtens arter – Nisaetus cirrhatus (Indien-art) og Nisaetus limnaeetus (Indokina m.m). Men cirrhatus er den nominelle art (type- eller referencearten), selv om limnaeetus også kan blive nævnt som sådan.
De to underarter må betragtes som dem der har haft størst påvirkning af slægtens artskompleks. Yderligere menes limnaeetus, at interagere med flere af slægtens underarter, hvorved den betragtes som en meta-art, og derved som havende større indflydelse på (nok) især Ø-gruppernes udseende.
Nisaetus cirrhatus (Lysbuget høgeørn / Changeable Hawk-eagle):
Nisaetus cirrhatus cirrhatus: Art status: nominel underart (typearten/referencearten)
Nisaetus cirrhatus ceylanensis: Art status; underart af Nisaetus cirrhatus
Nisaetus cirrhatus andamanensis: Art status; underart af Nisaetus cirrhatus
Nisaetus limnaeetus (Lysbuget høgeørn / Changeable Hawk-eagle):
Nisaetus limnaeetus limnaeetus: Art status: fuld art status, men en del af artskomplekset (kan nogle gange optræde som den nominelle underart). Slægtens mest udbredte underart.
Nisaetus limnaeetus vanheurni: (Art status: underart af Nisaetus limnaeetus
Nisaetus floris: Art status: – art-status (nyligt tildelt, og nu ikke en del af Lysbuget høgeørns gruppe af underarter).
Artskompleks
I biologien er et artskompleks en gruppe af nært beslægtede, der er så ens i udseende og andre træk, at forskellene mellem dem ofte kan være eller er uklare eller vanskelige at skelne fra hinanden – eller i bedste fald, kun adskiller sig på få/mindre områder. Taxaerne (grupperne) i komplekset kan være i stand til let at hybridisere (få fælles afkom) med hinanden.
Et artskompleks er i de fleste tilfælde en monofyletisk gruppe* med en fælles forfader. Kan ofte findes i forbindelse med underarter. Men der er undtagelser (dem kommer vi ikke ind på her).
Monofyletisk gruppe
En klade, er også kendt som en monofyletisk gruppe eller naturlig gruppe, er en gruppe af individer, der er monofyletiske – det vil sige nedstammende fra en fælles forfader og alle dens direkte efterkommere.
Den fælles forfader kan være et individ, en population eller en art.
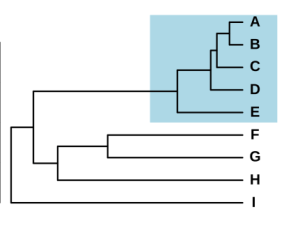
Et artskompleks danner typisk en monofyletisk gruppe, der har udviklet og spredt sig ret nylig, som det fremgår af de korte grene mellem arten A–E (blå boks) i dette fylogenetiske træ .
Overlap
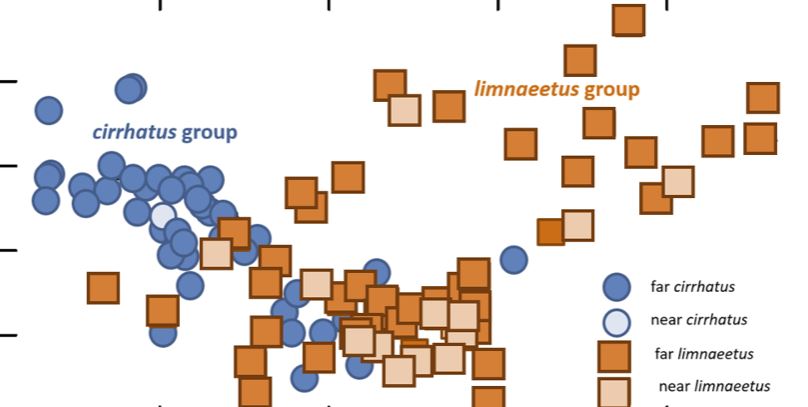
Selv om de enkelte arter er forholdsvis tæt knyttede til hver deres udbredelsesområde, forekommer der dog en overlapning, hvor især limniatus arten er kendt for at have en større kantmæssig udbredelse.
Her under ses angivelser for hvordan de to hovedgrupper chirrhatus og limnaetur træffes over deres udbredelsesområder. Hvor N. chirratus chirratus hovedsageligt træffes over Indien og Sri Lanka, og limniatus betydelig mere udbredt over Tailand og de forskellige Ø-områder.
Rovfugle er som gruppe ikke en biologisk eller taksonomisk gruppering, men en gruppe af fugle med ensartet udviklede egenskaber. Egenskaber som under den normale evolution, har været fordelagtige at erhverve under et liv som jæger eller et liv i ensartede biotoper
MEN:
- Hvorfor er alle kødædende fugle ikke rovfugle !
- Hvad afgør om en fugl, er en rovfugl ?
I 2019 kom en ny videnskabelig definition af hvad der skal til
for at en art indgår i – rovfuglegruppen: LÆS OM DEN HER

Taksonomisk placering af Lysbuget høgeørn
For at få overblik over hvor i rovfuglegruppen den Lysbugede høgeørn befinder sig, må vi også have et overblik over den fulde rovfuglegruppe Accipitriformes, hvor høgeørnene er placeret et godt stykke nede i det Taksonomiske klassificerings system.
Den taksonomiske orden kan ses i boksen over for, efterfølgende gennemgås kort de enkelte klassificeringer af; familie, underfamilie, slægt, art og underart.
Ordenen Accpitrifornmes (rovfugle orden)
Ordenen omfatter de fleste af de dagjagende rovfugle, herunder høge, ørne, glenter og den gamle verdens gribbe – men ikke falkene som omfattes af deres egen orden – Falkinoformes.
Ordenen udgøres i dag af 262 rovfugle arter fordelt i 75 slægter, som opdeles i 5 familier:
Accipitridae*: (ørne, glenter, høge (fx lysbuget høgeørn) og gl. verdens gribbe)
Pandionidae: (Fiskeørne)
Cathartidae: (ny verdens gribbe inkl. kondor)
Sagittaridae (sekretærfugl)
Cariamidae: Serimaer – ny familie iflg. rovfugledefinitionen af 2020, som kan ses HER.
Ordenen Accipitrifornmes (rovfugle orden) vist i undergrupper på vej mod Lysbuget høgeørn
Orden: Accipitriformes: Rovfugleorden
Familie Accipitridae: Familie med 13 underfamilier
Underfamilie: Aquilinae: Nævnes også som “støvle ørne” (bootede eagles) udgøres af 13 slægter.
Slæge: Nisaetus: er en de 13 slægter, og indeholder 10 arter, som vi vil komme ind på senere i teksten.
Art: Lysbuget høgeørn (Nisaetus cirrhatus) er en af de 10 arter i slægten Nisaetus. Den findes i 5 underarter som kan opdeles i to grupper efter om de har kam elles ej – ”signaturfjer”:
Familien Accipitridae
En af rovfugleordenens (Accipitriformes) 5 familier er Accipitridae familien (vist ovenfor), som er en familie med en stor variation af små til store rovfugle.
Familien indeholder 255 arter, som er opdelt i 70 slægter fordelt i 13 underfamilier (se boksen). De findes i alle dele af verden, undtagen Antarktis, og nogle oceaniske øgrupper. De lever i således mange forskellige biotoper, som ørkener, skove, græsarealer og
vådområder.
De lever af lige så forskelligartet byttedyr, som de lever i forskellige biotoper, lige fra insekter til mellemstore pattedyr, og nogle af ådsler og nogle få af frugt.
Mange velkendte fugle som f.eks. høge, ørne, glenter og gribbe fra den gamle verden er inkluderet i denne gruppe. Men Familien er meget forskelligartede i både form og størrelse idet de spænder lige fra den lille Dværgglente (Gampsonyx swainsonil) på 20-23 cm, til munkegrippen (Aegypius monachus) som måler hele 120 cm og har en vægt på op til 14 kg.
De fleste af slægterne deler en meget lignende morfologi, men mange af grupperne indeholder taxa , der er afviger meget fra hinanden. Disse er placeret i deres respektive position, mere af mangel på bedre beviser end noget andet. Det fylogenetiske
layout af accipitriderne har således altid været et grundlag til uenighed.
13 underfamilier
Elaninae – visse glenter
Gypaetinae – en af den gamle verdens to underfamilier af gribbe.
Perninae – honning-musvåger, Madagaskar-slangeørn) mf.
Circaetinae – slangeørne
Aegypiinae – gribbe i den gamle verden
Harpiinae – skovørne
Aquilinae* – støvleørne – “med Lysbuget høgeørn”
Harpaginae – Høge og visse glenter
Melieraxinae – melieraxine høge
Accipitrinae – accipitrinhøge
Circinae
Buteoninae – Våger, visse glenter og havørne

Underfamilien Aquilinae
Aquilinae er en af de 13 underfamilier hos Familien Accipitridae (se boksen ovenfor). Denne underfamilien betegnes også som “støvle ørn”, fordi de i modsætning til arterne i andre af gruppens underfamilier har fjer hele vejen op ad benene.
Underfamilien er opdelt i 10 slægter:
Stephanoaetus cornatus – Kronørn (crowned eagle)
Nisaetus* – Høgeørne, hvor Lysbuget høgeørn findes (Crested hawk-eagles group)
Spizaetus – høgeørne slægt med 4 arter
Lophotriorchis kienerii – Rødbuget høgeørn (Rufous-bellied eagle)
Polemaetus bellicosus – Kampørn
Ictinaetus – Sort ørn
Lophaetus – Lang-kammet ørn
Clanga – Skrigeørne; tre arter
Aquila – Kongeørn, Høgeørn, Kejserørn mf. Betegnes ofte som “de egentlige ørne”
Hieraaetus – meget nær beslægtet med Aquila, som er ”Type art” for denne slægt. Der hersker nogen uenighed om placeringen af disse arter.
Slægten Nisaetus (lysbuget høgeørn)
Nisaetus (Høgehøg-ørnegruppen) er en slægt i underfamilien Aquilinae (også kaldet Booted eagles/Støvleørne på grund af deres fjerbeklædte ben).
Nisaetus ørnene var tidligere placeret i slægten Spizaetus (høgeørne-gruppe) men offentliggørelsen af to molekylære fylogenetiske undersøgelser fra 2005 og 2007 vist, at de gamle verdens repræsentanter var tættere på slægten Ictinaetus (Sort ørn) end på den nye verdens Spizaetus arter. Dette resulterede i at Lysbuget høgeørn sammen med ni andre høge som alle findes i den gamle verden, blev flyttet til slægten Nisaetus – se boks her under.
De to art-grupper udgøres således nu af Spizaetus (som findes i den nye verden) og Nisaetus (som findes i den gamle verden). De udgør en høgeørne-gruppe levende i tropiske områder i Amerika, og en tilsvarende levende i tropiske områder i Asien.
Taksonomisk klassifikation
Klasse: Aves (alle fugle)
Orden: Accipitriformes (rovfugle gruppe)
Familie: Accipitriformes (Høge, ørne mf)
Underfamilie: Aquilinae (egentlige rovfugle)
Slægt: Nisaetus (høgeørne)
Art: Nisaetus cirrhatus (lysbuget høgeørn)
Oversigt over flytning fra gammel til ny slægt)
Figuren her under viser i et fylogenetisk træ slægtskabet mellem Spizatus og Nisaetus, og illustrerer det tættere slægtskab mellem fuglene i den etablerede Nisaetus slægt og den tilbageværende Spizaetus slægt.
Nisaetus og Kronørnen (Stephanoaetus coronatus) udgør sammen en klade*. Det samme gør Spizaetus sammen med Kastanieørnen ( Oroaetus isidori), da begge hver især kan føres tilbage tilbage til den samme fælles stamfader.

Slægt: Spizaetus de 4 tilbageværende arter af høgeørne.
Den tilbageværende slægt af Spizaetus, som er typiske høgeørne levende i de tropiske egne af Amerika.
Den resulterende og nuværende slægt Nisaetus – 10 arter af høgeørne.
Nisaetus alboniger (Sorthovedet høgeørn / Blyts Hawk-eagle)
Nisaetus bartelsi (Javahøgeørn / Javan Hawk-eagle)
Nisaetus cirrhatus (Lysbuget høgeørn / Changable Hawk-eagle)
Nisaetus floris (Flores høgeørn / Flores Hawk-eagle)
Nisaetus kelaarti (Legges høgeørn / Legges hawk-eagle)
Nisaetus philippensis (Filippinsk Høgeørn / Hawk-Eagle))
Nisaetus pinskeri (Pinskers Høgeørn / Pinskers Hawk-eagle)
Nisaetus nanus (Malajhøgeørn / Mountain Hawk-eagle)
Nisaetus nipalensis (Bjerghøgeørn / Mountain Hawk-Eagle)
Nisaetus lanceolatus (Sulawesihøgeørn / Sulawesi Hawk-Eagle
Den oprettede slægt af de flyttede Spizaetus (nu Nisaetus) arter. De er typiske høgeørne levende i store dele af det tropiske Asien.
Taksonomisk navnegivning
Lysbuget høgeørn (Nisaetus cirrhatus) findes i fem anerkendte underarter, hvor ”cirrhatus” (som nævnt) er den nominelle art, ”typearten” eller den art der refereres til. Hos Lysbuget høgeørn er cirrhatus altså den underart, der ligger som reference (eller til sammenligning) for andre underarter i denne art-gruppe.
Når en art eks.vis Lysbuget høgeørn ”Nisaetus cirrhatus” anerkendes som have en eller flere underarter, tildeles et tredje navn til artsnavnet, fx den Lysbuget Høgeørn som findes på Sri Lanka. Det tredje navn angiver så, at der er tale om en underart af ”Lysbuget høgeørn ”Nisaetus cirrhatus”” – den har fået artsnavnet ”ceylanesis”.
(Se mere om taksonomisk navnegivning HER)
Den Lysbugede høgeørn fra Sri Lankas fulde navn er så – Nisaetus cirrhatus ceylanesis (Lysbuget høgeørn fra Sri Lanka).
Den nominelle art (den der i første omgang lagde navn til selve arts-navnet), skal så også tillægges et tredje arts-navn, hvilket vil blive en tilføjelse af dens navn i Nisaetus slægten ”cirrhatus”. Dens fulde navn vil så være – Nisaetus cirrhatus cirrhatus. Dette viser så, at der er tale om ”Indien-arten”.
Se nærmere i gruppens underarter her under.
Slægten Nisaetus – 10 arter med underarter
Slægtnavn: Nisaetus
Artsnavn: Lysbuget høgeørn (Nisaetus cirrhatus – den navnebærende typeart) 5 underarter;
- Nisaetus cirrhatus cirrhatus – Den nominelle art fra Indien
- Nisaetus cirrhatus ceylanesis – Sri Lanka underarten (endemisk)
- Nisaetus cirrhatus limnaeetus – Nepal, nordøstlig Indien, Burma og Malaisia – ingen kam
- Nisaetus cirrhatus andamanensis – Andaman-ørene – kun lille varierende kam 0-5 cm.
- Nisaetus cirrhatus vanheurni – Simeulue-øerne – ingen kam
Flores Høgeørn – Nisaetus floris (Flores Hawk-Eagle)
Bjerghøgeørn – Nisaetus nipalensis (Mountain Hawk-Eagle ) (2 underarter);
Nisaetus nipalensis nipalensis
Nisaetus nipalensis orientalis
Legge’s Hawk-Eagle – Nisaetus kelaarti (Legge’s Hawk-Eagle)
Sorthovedet Høgeørn – Nisaetus alboniger (Blyth’s Hawk-Eagle)
Javahøgeørn – Nisaetus bartelsi (Javan Hawk-Eagle)
Sulawesihøgeørn – Nisaetus lanceolatus (Sulawesi Hawk-Eagle)
Filippinsk Høgeørn – Nisaetus philippensis (Philippine Hawk-Eagle)
Pinsker’s Hawk-Eagle – Nisaetus pinskeri (Pinsker’s Hawk-Eagle)
Malajhøgeørn – Nisaetus nanus (Mountain Hawk-Eagle ) (2 underarter
Nisaetus nanus nanus
Nisaetus nanus stresemanni




Kladiogram (fylogenetisk stamtræ)
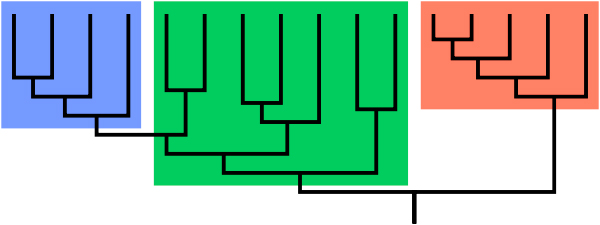
Skematisk oversigt over et kladiogram (stamtræ) af en biologisk gruppe. Den sidste fælles forfader angives ved den lodrette linjestamme i bunden. Sammenlagt udgør de tre farver en klade, idet de kan ledes tilbage til en fælles stamfader.
Set opdelt i tre grupper, udgør den blå og røde undergruppe to klader; hver viser sin fælles stamfader i bunden af undergruppe-grenen. Den grønne undergruppe er nu ikke en klade; det er en parafyletisk gruppe, fordi den udelukker den blå gren, som er nedstammet fra den samme fælles forfader. Samles den grønne og blå undergrupper danner de tilsammen en klade, fordi alle den fælles forfaders afkom er samlet.
Betragtes den grønne og den røde særskildt, har vi igen en parafyletisk gruppe, fordi den fælles stamfader også er forfader til blå. En klades stamfader SKAL indeholde alle hans efterkommere.
Navnesystematikken
Forstå det sammensatte navns betydning
Hvordan angives og forstås fuglenes navne. Nogle fugle angives ved to navne andre ved brug af tre ?
I biologisk klassificering refererer udtrykket underart til en af to eller flere populationer af en art, der lever i forskellige områder af artens udbredelse, og som på en eller flere måder varierer fra hinanden. En art anerkendes enten som at have ingen underarter overhovedet eller mindst to.
Arten er systematikkens basale enhed, den der specifikt skal navnegives, så den enkelte art ikke kan forveksles med andre. I teksten her er navnet ”kongeørn = chrysatos” (den latinske betegnelse), specifikt, og kan ikke forveksles med et andet fuglenavn. Dette navn staves altid på korrekt vis med et lille bogstav.
Her ud over skal arten knyttes til en ”slægt”, som altid staves med stort – i dette tilfælde ”Aquila”
Det videnskabelige navn hos en slægt som består af en enkelt fugleart, betegnes således ved to navne – og for kongeørnen ”Aquila chrysaetos” = slægt og art.
Men, hvis der findes og anerkendes yderligere en art af kongeørnen, og der så findes to arter (eller flere) i samme art-gruppe, må endnu et tredje navn tilføjes. Herefter har alle arter i slægten tre navne. Den først anerkendte art af kongeørn som har det videnskabelige navn ”Aquila chrysaetos”, skal så tilføjes et tredje navn.
Tre navn bliver således efterfølgende permanent for alle arter i Kongeørn art-gruppen. Gruppes første art Kongeørnen ”Aquila chrysaetos”, betegnes så Aquila chrysaetos chrysaetos, for det er det tredje navn, som fastslår, at det lige netop er den Europæiske Kongeørn (chrysaetos) der er tale om. Den næste art i slægten betegnes Aquila chrysaetos japonica, hvilket fastslår, at det er den Japanske variant af Kongeørn (japonica) der er tale om = slægtnavn, artsnavn og den specifikke underart ”japonica eller chysaetos.
En tredje underart i slægten kan så hedde ” Aquila chrysaetos homeyeri” det tredje navn ændres, så det angives, at det er den Iberiske variant af kongeørn, og således efterfølgende. Alle arter i gruppen kommer til at hedde ” Aquila chrysaetos” med et efterfølgende specifik underarts navn.
Disse tre arter er så underarter i art-gruppen ”Aquila chrysaetos” (se boksen – Slægten Aqulia).
Typearten
*Typearten” repræsenterer referencearten (eller den nominelle art) og danner dermed “definitionen” for et bestemt slægtsnavn.
For kongeørnen er typearten “Aquila chrysaetos” (Europæisk kongeørn), som således er reference arten, som gerne vises øverst i gruppen.
Den nominelle art, er den navnebærende variant af en slægt eller underslægt.
Typearten knytter permanent et formelt navn (det generiske navn) til en slægt (eller en art i en slægt med flere arter), ved kun at angive én art inden for den pågældende slægt, som slægtsnavnet er permanent knyttet til (dvs. slægten skal omfatte den art, hvis den skal bære navnet). Det er altid den først anerkendte og beskrevne art.
Hybrid
Er afkom af individer af forskellige underarter, racer, arter eller slægter.
Som oftest vil hybrid-afkom ikke være forplantningsdygtige, men hvis forældreparret slægtsmæssigt ligger meget tæt på hinanden, kan det godt resultere i at hybriderne kan producere afkom.
Hybride zoner.
En hybridzone er et område, hvor to arter eller underarter begge træffes. De kan fremvise og indeholde en eller begge rene fænotyper plus førstegenerations- og tilbagekryds-hybrider.
Fænotype; det man ser / individets udseende
ANBRINGES VED OVERLAP ELLER HYBRID